


**The AGA grants EECG Research Awards each year to graduate and post-doctoral researchers who are at a critical point in their research, where additional funds would allow them to conclude their research project and prepare it for publication. EECG awardees also get the opportunity to hone their science communication and write three posts over their grant tenure for the AGA Blog. In the first in the series, our EECG awardees write about their research and their interests as an ’embarkation’.

About the Author: Dr. Rachel Moran is a Postdoctoral Associate in Dr. Suzanne McGaugh’s lab at the University of Minnesota and a 2021 EECG Awardee. Her research integrates behavioral and genomic approaches to study sexual selection, local adaptation, and speciation in fishes. To learn more about Rachel’s work you can visit her website and follow her on twitter @Rachel_L_Moran.
A major goal in evolutionary biology is to understand how lineages diverge in the face of gene flow. Gene flow and recombination have traditionally been thought of as homogenizing forces, acting to break up co-adapted gene complexes that differ between diverging lineages (Turelli et al. 2001). Within the past decade, an increase in the availability of genomic data has made it possible to look for signatures of recent and ancient introgression in natural populations (reviewed in Abbott et al. 2016, Taylor & Larson 2019). Surprisingly, this work has demonstrated that gene flow and hybridization (i.e., gene flow that occurs between members of two different species) often play an important role in species diversification.
Reinforcement is one type of speciation with gene flow that can act to finalize reproductive isolation in sympatry after secondary contact. Reinforcement occurs when natural selection directly favors the evolution of prezygotic barriers (e.g., behavioral isolation) in response to the presence of postzygotic barriers (e.g., genetic incompatibilities) (Butlin 1987, Howard 1993). In this manner, hybridization can actually promote the process of speciation.
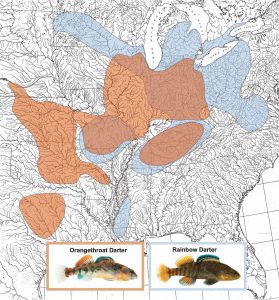
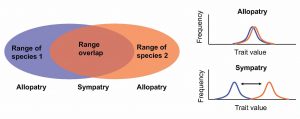
My research examines the genetic basis of reinforcement in a diverse group of North American stream fishes called darters. Much of my work has focused on two wide-ranging species that overlap in several areas of sympatry, the orangethroat darter (Etheostoma spectabile) and the rainbow darter (Etheostoma caeruleum) (Figure 1). I found that males of these species have strong mating preferences for conspecific females (versus females from the other species), but only in populations where they naturally co-occur in sympatry. If fish from allopatric populations of both species – which have never encountered the other species in the wild – are brought together in the lab, males do not discriminate between females of the two species (Moran & Fuller 2018). This pattern of enhanced behavioral isolation in sympatry compared to allopatry is a classic signature of reinforcement, and is also referred to as reproductive character displacement (Figure 2). (As an aside, although the terminology used to describe character displacement and reinforcement in the literature has historically been inconsistent, the current consensus appears to be that a pattern of reproductive character displacement can come about through many underlying mechanisms/selective pressures, and reinforcement/selection against hybridization is just one of those possible mechanisms.)
My work in darters has also shown that reinforcement drives the pattern of reproductive character displacement in this system. Hybrids suffer from negative fitness consequences in the form of skewed sex ratios in F1s and heightened mortality in backcrosses (Moran et al. 2018). In Moran et al. (2018), we found that F1 hybrid crosses between orangethroat and rainbow darters result in highly male-skewed sex ratios in both cross directions. Parental crosses do not deviate from a 1:1 male to female sex ratio. Interestingly, females do not appear to be missing from hybrid clutches, as F1 and parental crosses do not differ in total clutch size or number of offspring that survived it to adulthood. This suggests that F1 individuals that are genetically female may appear phenotypically as male, potentially due to genetic incompatibilities at the sex determining region (SDR). Such an incompatibility could act as a postzygotic barrier between species.
This finding is intriguing because regions with low recombination, such as chromosomal inversions and SDRs, have been implicated in reinforcement in other systems (e.g., Ficedula flycatchers: Sæther et al. 2007; Drosophila: Noor et al. 2001). Theory suggests that regions of reduced recombination could facilitate the evolution of reinforcement by maintaining linkage disequilibrium between loci underlying prezygotic and postzygotic barriers (reviewed in Ortiz-Barrientos et al. 2009). However, investigations into the genetic basis of reinforcement have been rare. Additional empirical work is needed to resolve whether certain aspects of genomic architecture are more likely to resist introgression and drive speciation via reinforcement.
As part of a project supported by an AGA EECG Research Award, I am currently working towards confirming whether genetic incompatibilities in the SDR are present between orangethroat and rainbow darters and might in turn promote reinforcement. I am using whole genome sequencing to: (1) identify the SDR in orangethroat and rainbow darters, (2) ask whether the SDR differ in physical location and/or exhibit coding changes between species, and (3) determine whether differences in the SDR are present between sympatric and allopatric populations within species. Genomic analyses will be paired with crosses between allopatric orangethroat and rainbow darter populations and analysis of resulting F1 hybrid sex ratios. If sex ratios are skewed in F1 crosses between allopatric populations, similar to what was previously observed in F1 crosses between sympatric populations, this would suggest that the associated genetic incompatibilities were present before secondary contact in sympatry between orangethroat and rainbow darters. Furthermore, such a finding would imply that genetic incompatibilities in the SDR may have favored the evolution of behavioral isolation in sympatry between these species (i.e., through reinforcement). Ultimately, this work could provide insight into the mechanisms that promote diversification between hybridizing species.
References
Butlin, R. (1987). Speciation by reinforcement. Trends in Ecology & Evolution, 2(1), 8-13.