


**The AGA grants EECG Research Awards each year to graduate and post-doctoral researchers who are at a critical point in their research, where additional funds would allow them to conclude their research project and prepare it for publication. EECG awardees also get the opportunity to hone their science communication and write three posts over their grant tenure for the AGA Blog. In the first in the series, our EECG awardees write about their research and their interests as an ’embarkation’.
 About the Author: I am a fourth year NERC-funded PhD student at The University of Sheffield, supervised by Dr Alison Wright. Broadly, I am fascinated by the complex genomic and transcriptional architecture underlying biodiversity. During my PhD I have been studying this by focussing on sex differences. This is because sexual dimorphismis, in many species, the most striking example of intraspecific biodiversity there is. The sexes are often different not only in their basic reproductive traits, but also in their morphology and behaviour. However, males and females of the same species share the majority of their genomes. My research aims to uncover how these sex differences arise, by focussing on both the role of regulatory evolution across the shared part of the genome as well as the role of sex chromosomes in the evolution of sexual dimorphism. Twitter handle: @TheaFRogers.
About the Author: I am a fourth year NERC-funded PhD student at The University of Sheffield, supervised by Dr Alison Wright. Broadly, I am fascinated by the complex genomic and transcriptional architecture underlying biodiversity. During my PhD I have been studying this by focussing on sex differences. This is because sexual dimorphismis, in many species, the most striking example of intraspecific biodiversity there is. The sexes are often different not only in their basic reproductive traits, but also in their morphology and behaviour. However, males and females of the same species share the majority of their genomes. My research aims to uncover how these sex differences arise, by focussing on both the role of regulatory evolution across the shared part of the genome as well as the role of sex chromosomes in the evolution of sexual dimorphism. Twitter handle: @TheaFRogers.
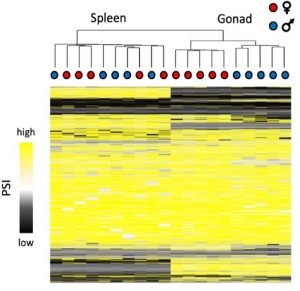
Because the sexes share the majority of their genomes, it is widely assumed that sex differences in transcriptional regulatory mechanisms facilitate the decoupling of male and female phenotypes, resulting in sex-specific adaptation (Connallon & Knowles, 2005; Innocenti & Morrow, 2010; Mank, 2017). Sex‐specific expression allows selection to act independently on males and females and therefore the genome can evolve in a sex-specific manner. So far, research has mainly focussed on the role of genes with sex-biased expression levels in the evolution of sexual dimorphism. However, gene expression level is only oneelement of a complex transcriptional framework and other gene regulatory mechanisms have been overlooked. In addition to expressing the same gene at different levels, the sexes can express different protein isoforms. This is accomplished by sex-biased alternative splicing, where different exons are spliced or shuffled in males and females to create distinct sex-specific sequences (Blekhman et al., 2010; Nilsen & Graveley, 2010). There is growing evidence, including my own,that alternative splicing varies across species, sexes and tissues (Fig. 1), and has important phenotypic consequences (Rogers et al., 2020; Schütt & Nöthiger, 2000; McIntyre et al., 2006). My previous work, published in Molecular Biology and Evolutionhas also shown that when genes are subject to pleiotropic constraints, the evolution of sex-specific isoforms may offer a more viable mechanism than changes in expression level to achieve sex-specific functions (Rogers et al., 2020). However, studies of alternative splicing, including mine, primarily assess transcription in adults, but many adult dimorphisms are established during embryogenesis and it is simply not appropriate to only study them outside of their developmental context. Additionally, our previous work only focussed on three species. My future work will therefore assess transcriptional regulation across multiple developmental stages and additional species, which my recent EEECG AGA award will help support.
References