



About the Author:
Hossein Madhani is a second-year Ph.D. student in Ecology and Evolutionary Biology at the University of Nevada, Las Vegas, where he is a member of Dr. Elizabeth Stacy’s lab. He received his B.Sc and M.Sc in Plant Biology from the University of Tehran, Iran, where he studied the molecular phylogeny and taxonomy of carnations (Caryophylleae tribe) (Madhani et al., 2018), and the dynamics of diversification in the same group of flowering plants (Madhani et al., in prep)
The appearance of hybrid incompatibilities (HI) as a sign of divergence between populations is a pivotal stage in the continuum of speciation (Coyne et al., 2004). Hybrid necrosis (HN) is a common type of HI in plants caused by detrimental interactions between components of the immune system (disease-resistance genes). These detrimental interactions are consistent with the Bateson-Dobzhansky-Muller model in which alleles that are coadapted within populations are incompatible between populations (Bomblies et al., 2007; Bomblies & Weigel, 2007; Li & Weigel, 2021; Wan et al., 2021). Studies show that negative epistatic interactions between alleles of resistance genes, primarily those encoding nucleotide-binding and leucine-rich repeat (NLR) receptors, are the primary cause of HN (Wan et al., 2021). Evidence for HN derives predominantly from model species or crops, however, and little is known about the role of HN in speciation in nature. Because they capture a range of incomplete isolating barriers, ongoing adaptive radiations are ideal for investigating the evolution of isolating barriers – through HN or other mechanisms – during speciation.
Hawaiian Metrosideros: a model of island adaptive radiation
Hawaiian Metrosideros Banks ex Gaertn, belonging to the Myrtaceae family, is a highly diverse woody species complex that dominates the islands’ native forests wherever rainfall exceeds 500 mm annually (Stemmermann & Ihsle, 1993). Over the course of its 3-4 million-year history on the islands (Dupuis et al., 2019; Percy et al., 2008; Wright et al., 2000), this single-origin group has evolved into more than 20 taxa or morphotypes distinguished through vegetative characters (Dawson & Stemmermann, 1990; E. A. Stacy & Sakishima, 2019). As a group, Metrosideros demonstrates adaptation to an extraordinary range of environments, including a 2,500-meter elevation gradient, an annual rainfall range of over 8,500 mm, an annual temperature variation of 14 C, and a 4-million-year substrate-age gradient (Dawson & Stemmermann, 1990; Giambelluca et al., 2013; Kitayama et al., 1995; Kitayama & Mueller-Dombois, 1992). All taxa showcase an incredible capacity for gene flow, enabled by bird- and insect-mediated pollen dispersal (Koch & Sahli, 2013) and wind-assisted seed dispersal (Drake, 1992). Within continuous forest on each island, the taxa of Hawaiian Metrosideros are nonrandomly distributed across a range of environments, from new lava flows and mature forests to deserts, bogs, subalpine zones, riparian zones, and windy cliffs. These taxa exhibit remarkable heritable phenotypic variation (Stacy & Johnson, 2021), ranging from the mat plants that bloom at a height of a few inches (M. polymorpha race prostrata) to the 40-meter tall trees (M. polymorpha var. polymorpha), particularly in the richest soils of Hawaii Island where trees commonly live over 650 years (Hart, 2010).

Evidence of differential local adaptation to various environments, partial reproductive isolating barriers (Rhoades, 2012; Stacy et al., 2017), and a wide range of genetic distances among the taxa indicate that Hawaiian Metrosideros is an emergent adaptive radiation. This feature implies that the group captures multiple stages of the speciation process, making it a valuable model for understanding speciation. The partial isolating barriers observed between taxa include HN in F1 offspring of several hybrid crosses. For example, ~8-20% of the F1 seedlings from controlled crosses between Metrosideros polymorpha var. newellii (newellii) and M. polymorpha var. glaberrima (glaberrima) exhibit reduced growth rate and dwarfism, tissue necrosis, chlorosis, and yellowing (Fig. 1) identical to the autoimmune-associated HN phenotypes described in model systems (Bomblies et al., 2007; Wan et al., 2021). These two sympatric taxa, newellii and glaberrima, have diverged recently through disruptive selection across a sharp forest-riparian ecotone in the presence of gene flow (Choi et al., 2020; Ekar et al., 2019). We posit that adaptation of Hawaiian Metrosideros to contrasting environments exposes these trees to different pathogens, and that the resulting divergent selection on resistance genes results in HN in hybrid offspring. This assertion is supported by the theory of local adaptation, which postulates that organisms adapt to their local environments in ways that maximize their fitness, including the evolution of disease resistance in response to the presence of pathogens (Kawecki & Ebert, 2004).
The primary aim of our research is to map the diversity of NLRs and uncover patterns of selection and introgression on these genes within the Hawaiian Metrosideros radiation. To this end, we are leveraging population-level resequencing data from 191 individuals across 12 key populations/taxa by the use of traditional population genetic statistics along with machine learning. Additionally, we are implementing RNA sequencing and metabolite profiling techniques to contrast normal and necrotic phenotypes. This multifaceted approach will not only help us to achieve our core objectives but will also aid in identifying candidate regions associated with the HN phenotype.
New insights
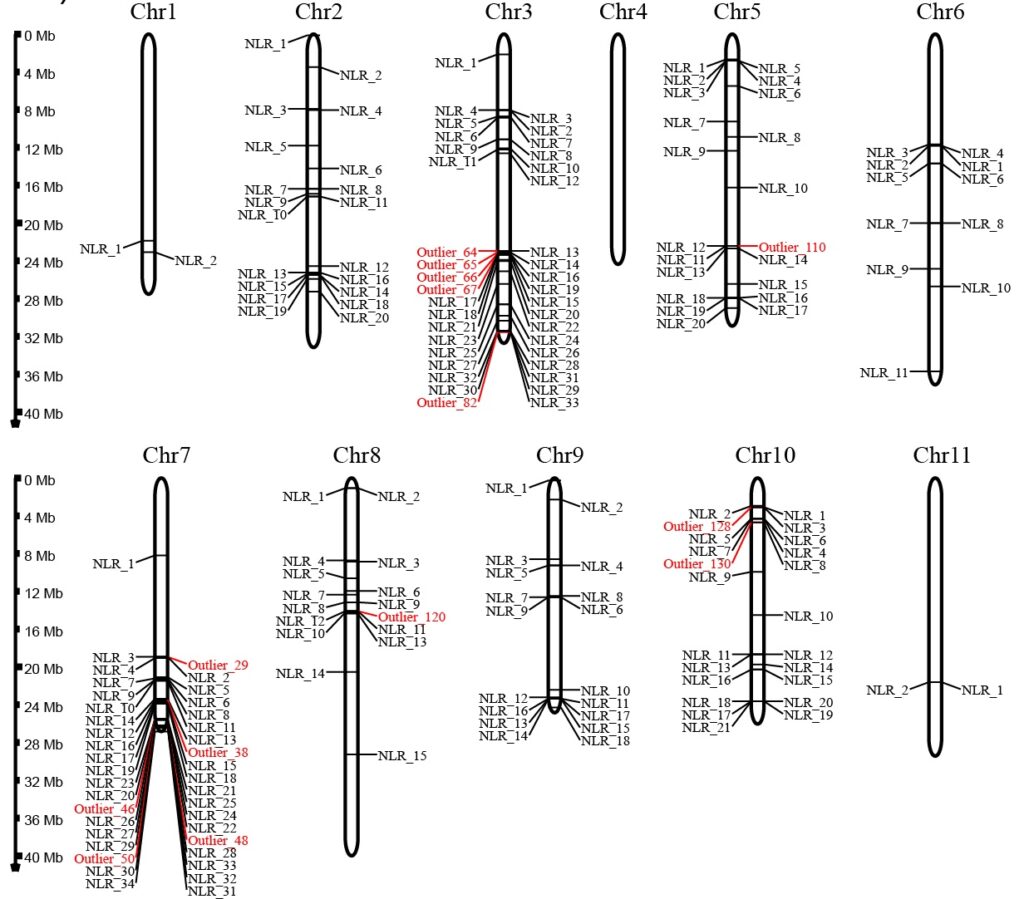
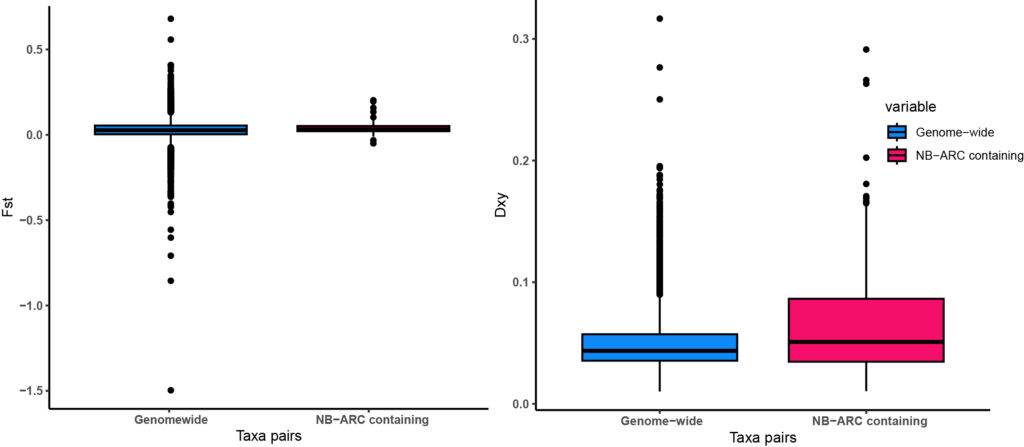
We have identified a final set of 180 putative NLRs in Hawaiian Metrosideros, which were subsequently used for variant calling and population genomics analyses. The findings revealed a subset of NLRs on chromosomes 3, 7, and 10, distinguished by high nucleotide diversity across the radiation (Fig 2). These NLRs present themselves as potential HN genes. The population-pairwise Fst values for NLRs align with genome-wide Fst values observed earlier (Choi et al., 2021). However, the heightened Fst and Dxy values for NLRs between glaberrima and newellii (Fig 3), in conjunction with D statistics observed for the same pair (not illustrated here), support the involvement of NLRs in the HN phenotype observed in hybrids.
The results of this study will uncover the patterns of selection on NLR genes across the Hawaiian Metrosideros radiation, and provide insight into the genetic architecture and putative loci associated with HN in this ongoing adaptive radiation. These results will improve our understanding of the role of selection on resistance genes in the evolution of reproductive barriers in nature.