


**The AGA grants EECG Research Awards each year to graduate students and post-doctoral researchers who are at a critical point in their research, where additional funds would allow them to conclude their research project and prepare it for publication. EECG awardees also get the opportunity to hone their science communication and write posts over their grant tenure for the AGA Blog. In the wrap-up to the series, awardees talk about their award and research in their ‘epilogue’.**

About the Blog Author: Kathryn (Katie) Uckele is a PhD candidate at the University of Nevada Reno where she investigates the genetic and morphological outcomes of hybridization among long-lived conifers in western Nevada. Follow Katie on Twitter @katieuckele or via her website.
The study of hybrid zones advanced enormously in recent years due to next-generation sequencing data and its applicability to non-model organisms. Suddenly, genomic approaches could be applied to natural hybrid zones cheaply and efficiently, providing greater insight into genomic patterns of admixture and divergence, the strength and mechanism of reproductive barriers, and the genetic architecture of complex phenotypes (Gompert et al., 2017).
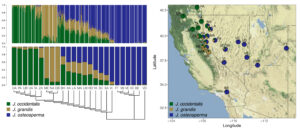
Hybrid zones often form along environmental gradients, where sharp ecological transitions promote local adaptation and isolation by environment. For example, altitudinal gradients are characterized by rapid changes in temperature, moisture, seasonality, geology, and sunlight, which are often, though not always, highly effective barriers to gene flow. Along the border of California and Nevada, three juniper species hybridize along the transition from the montane forests of the Sierra Nevada to the high desert of the Great Basin (Figure 1). This transition is marked by dramatic changes in seasonality of precipitation and temperature, which are accompanied by equally dramatic changes in the compositions of floral communities. These juniper species occupy distinct altitudinal zones and hybridize at intermediate elevations between those zones, forming hybrids with ancestry from all three parental species.

To understand patterns of admixture across the hybrid zone, my collaborators and I collected leaves from 326 individuals of Juniperus grandis, J. occidentalis, J. osteosperma, and putative hybrids across Oregon, California, Nevada, and Utah, and used a reduced-representation sequencing approach (ddRADseq) to generate data at 9,125 loci. A Bayesian model was used to elucidate hybrid ancestries and hybrid zone structure (Figure 2). Ancestry formed a discernible cline from higher to lower elevations (i.e., west to east). At the cline’s point of inflection, F1 and backcrossed hybrids occurred sympatrically. West (upslope) of the inflection point, the transition from hybrid to parental ancestry was steep, whereas east (downslope) of the inflection point, the transition was more gradual, suggesting asymmetrical gene flow from west to east. F2 and later-generation hybrids were extremely rare, but backcrossing was common. The geographic area occupied by hybrids was associated with climatic variation that was predominantly intermediate to that of the parental species, though a subset of hybrid populations existed in particularly dry and hot climates.
In addition to resolving patterns of admixture and hybrid zone structure, my collaborators and I wanted to understand how leaf terpenoid chemistry varied across the hybrid zone in response to genetic and environmental variation. Conifers, and junipers in particular, exhibit diverse terpenoid profiles that are important for herbivore resistance and signaling of natural enemies (Keeling and Bohlmann, 2006). Hybrid admixture can have disruptive effects on herbivore resistance, but can also facilitate the diversification of plant secondary chemistry (Cheng et al., 2011). In 2018, I received funding through AGA’s EECG award to conduct metabolomic analyses of the parental and hybrid individuals and began the long-awaited journey from silica-dried leaf material to gas chromatography column. I made a number of rookie mistakes, the worst of which was forgetting to add an internal standard to over 300 chemical extractions, which meant re-extracting every single sample. I may have given up, if not for the support of students and chemists at the University of Nevada Reno who donated long hours at the lab and designed a new extraction protocol. By late 2020, my collaborators had quantified the concentrations of 163 terpenoid compounds in over 300 juniper individuals across the hybrid zone. A PCA of this data revealed that while terpenoid variation was partitioned by species, a larger proportion of the variation was due to intraspecific differences in concentrations. For example, a number of hybrids exhibited transgressive concentrations which exceeded the range of the parentals. Surprisingly, the geographic position of populations was as important as genetic variation in explaining terpenoid variation across parental individuals and hybrids, and variation in moisture and temperature was comparatively insignificant. I hope to share these results and more in a manuscript that I am preparing to submit to the Journal of Heredity later this month.
References