



Brendan J. Pinto (he/him) received a PhD in August (2020) in the Gamble Lab. Currently he is a research associate of zoology at the Milwaukee Public Museum and a postdoctoral research fellow at the University of Texas at Austin in the Kirkpatrick Lab where they aim, among many other things, to disentangle the selective forces governing sex chromosome turnover in animals. For updates on past and future work, follow him on Twitter @DrPintoThe2nd.
Sexual reproduction is dependent on a population’s ability to bin some individuals as female (carrying eggs) and others as male (carrying sperm). Although sexual reproduction is ancestral in vertebrates, for instance, the ways in which different species go about determining sex (at the molecular level) varies greatly. In amniotes, specifically, there are two common ways for determining sex: temperature-dependent sex determination (TSD) and genetic sex determination (GSD).
TSD occurs when temperature at a critical point during incubation determines sex which has evolved in crocodilians, some turtles and squamates (i.e., lizards, snakes, and amphisbaenians), and the tuatara. Conversely, GSD occurs when the presence/absence of a particular genomic region determines sex. The latter can further be broken down into two ‘sub-types’: i) male heterogamety (commonly referred to as XX/XY systems), where the male possesses the sex-limited (Y) region of the genome, present in placental mammals, including humans, and has evolved independently many times including pleurodont lizards and some turtles and snakes; and ii) female heterogamety (or ZZ/ZW systems), where the female possesses the sex-limited (W) region of the genome, observed in birds and has evolved independently many times in squamates, including all ‘advanced’ snakes and varanid lizards. Although we have identified some macro-scale patterns across vertebrates, we have barely begun to scratch the surface of how many transitions there have been (even just within vertebrates!) and cataloging the patterns in vertebrates is an important first step to studying the processes involved in sex chromosome evolution more broadly.
One zealous avenue of sex chromosome research is identifying the sex chromosome linkage groups (or chunks of the genome that are conserved across large phylogenetic distances) within and amongst groups with high rates of sex chromosome turnover. Indeed, one hypothesis involving sex chromosome turnovers suggests that some chromosomes will be recruited repeatedly and preferentially over others (Graves & Peichel, 2010; O’Meally et al. 2012). This could be due to a few possibilities, but namely that some chromosomes may be betterat becoming a sex chromosome than others (in that certain linkage groups may maintain important genes in the sex determination cascade). To test this, we need sex chromosome linkage information across many independent transitions (i.e., from XX/XY to ZZ/ZW, or XX/XY1to XX/XY2, or any one of the myriad other possible permutations of transitions between and among GSD and TSD). To test these kinds of important hypotheses, we need exemplar model systems that have many, many transitions to and from various systems, with various levels of conservation and degeneration.
Squamate reptiles are particularly great models to study sex chromosome evolution. This is because groups of squamates rarely maintain ancient, conserved sex chromosome systems—unlike mammals and birds. This provides many natural experiments to study the ways in which sex chromosome systems transition and how they do, or don’t, degenerate. Indeed, just within squamates some groups are prone to frequent transitions between sex determination systems, while others aren’t. For instance, in geckos (>2,000 spp.), more than 30 transitions between TSD, XY, and ZW systems have been estimated to date, while in pleurodonts (~1,200 spp.), there has only been one transition. However, as many squamate groups lack such data, even within well-known groups such as skinks and their relatives (a.k.a. the scincomorphs), thus our ability to understand of how sex chromosomes evolve is severely hampered at present.
Recently, work in scincomorphs (which include skinks, cordylids, plated lizards, and night lizards) has provided an interesting case study in this debate. In our recent paper published in the Journal of Heredity, we identified a new ZW system within night lizards (Nielsen et al. 2020). This report of a ZZ/ZW system in Xantusia henshawiwas the first report of a sex chromosome system within scincomorphs, outside of skinks proper (the family Scincidae). Interestingly, we not only identified a ZZ/ZW system within Xantusia henshawi,butwe were also able to identify the linkage group within the genome that determines sex—a region that has never before been identified as a sex chromosome in any amniote. Unfortunately, these findings were limited in their utility to study macroevolutionary patterns of sex chromosome evolution, by the fact that we knew almost nothing about the sex chromosomes in most scincomorphs (see Figure 1; Nielsen et al. 2020). Coincidentally, it so happens that these findings were just the beginning of what has turned 2020 into the proverbial ‘year of the scincomorphs’!
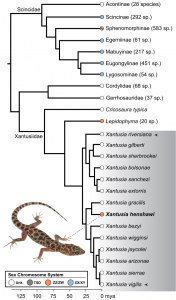
Since May, complimentary work by two other research groups has provided better context to begin asking questions about the evolution sex determination systems within the scincomorphs. First, independent research by groups at the National Autonomous University of Mexico (UNAM) and Charles University in Prague (Czech Republic) descended upon the hypothesis that temperature-dependent sex determination isn’t as prevalent in skinks as once believed and that, in fact, most skinks share a conserved XX/XY sex chromosome system—with a different linkage group than what we found in Xantusia (Cornejo-Paramo et al. 2020; Kostmann et al. 2020)! The plot thickens further when considering another recent study by researchers based at Charles University in Prague who were able to identify a likely ZZ/ZW system in the sister genus to Xantusia—Lepidophyma (Kratochvíl et al. 2020). These additional findings, as they relate to sex chromosome evolution in scincomorphs, are summarized here as Figure 1. Excitingly, in a single year, our understanding of sex chromosome evolution in scincomorphs has increased markedly. This will allow us to test more informed hypotheses about how sex chromosomes have evolved within this group. Specifically, we can now be certain that there is at least one XX/XY to ZZ/ZW transition in skinks (within the subfamily Sphenomorphinae; Patawang et al. 2017). Further, only a modest dataset is needed to determine if the ZZ/ZW systems in Xantusiaand Lepidophymaare conserved from a common ancestor, or independently-derived systems. Needless to say, any future research in scincomorph sex chromosomes has its work largely cut out for it!
I’d like to thank my collaborators on the Journal of Hereditypaper associated with this blog post: Andira Guzmán-Méndez, Tony Gamble, and especially to Stuart Nielsen for providing an original copy of Figure 3 from the manuscript. Personal copies of the associated manuscript is available from the author upon request:
Other References