



About the Blog Author:
Jade Mellor (she/her) is a PhD student in Dr. Greer Dolby’s lab focusing on evolutionary and conservation genomics in desert gopher tortoises. Previously, she has completed a B.S. in Biology at the College of William & Mary, and an MS in Biology from Clemson University. She wrote this blog post as part of Dr. Stacy Krueger-Hadfield’s Ecological Genetics course at the University of Alabama at Birmingham. Follow Jade on twitter @njademellor.
It is the 1990’s, and the Florida panther (Puma concolor coryi, Figure 1) is an endangered subspecies whose historical range once spread into the land east of the Mississippi but now is constrained only to southern Florida. Puma concolor coryi experienced a large bottleneck in the 1990s, when the population was estimated to be just 30 individuals (Hedrick, 1995). Due to their unfortunate situation, they are a great study system for understanding the negative effects of inbreeding, genetic drift, and genetic isolation (Ochoa et al., 2022).

Florida panthers had become genetically (and geographically) isolated from other panther populations. What had once been a wide range was restricted to three regions in south Florida. This physical isolation leads to genetic isolation, which results in inbreeding. Florida panthers have small population sizes and little mate choice. High amounts of inbreeding leads to inefficient purifying selection, where high homozygosity leads to deleterious (bad) traits such as heart defects and high parasite loads becoming fixed in the population. Conservationists were concerned about the future of the Florida panther.
This is because the genetic health of a population (or species for that matter), is an important indicator used by conservationists and wildlife agencies to determine their management approach. Genetic health can be used to assess how robust a population is, and if inbreeding is occurring (Ochoa et al., 2022).
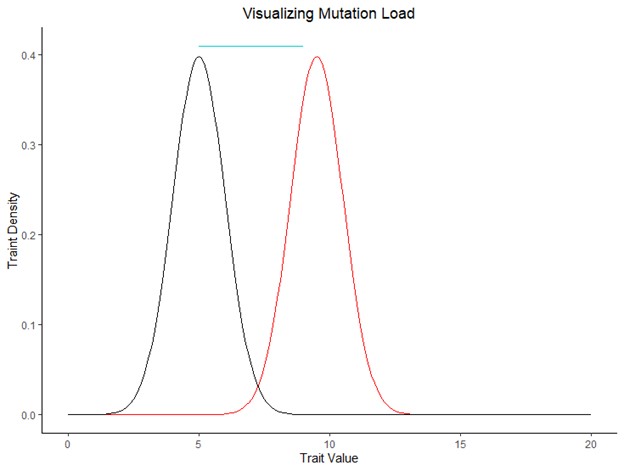
One metric of genetic health is mutation load, or the reduction in fitness caused by a widening gap between the ideal phenotype and the average phenotype in the population (Figure 2). This widening of the gap is caused when deleterious alleles are widely distributed in the population. There are three main ways deleterious alleles can become widely distributed (Hendricks, et al. 2018). It first begins with genetic isolation; populations do not share a gene pool with other populations. This leads to less mating options due to lack of connectivity and a smaller population size overall. Fewer mating options means less diversity in the gene pool, resulting in inefficient purifying selection. Deleterious alleles are unable to be removed from the gene pool through selection, and this can result in high homozygosity. There is no other option and so the deleterious alleles cannot be purged.
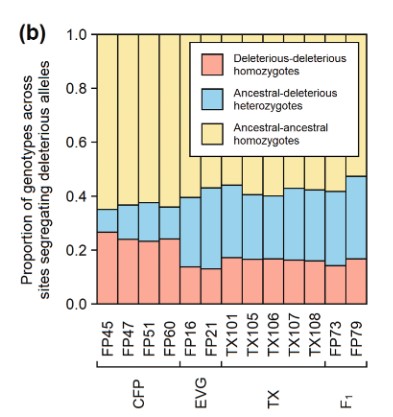
A solution to this is to outbreed one population by introducing individuals from another population in order to add genetic diversity. Conservationists did this in 1995 by introducing eight Texas panthers (P. c. stanleyana) (Ochoa et al., 2022). These Texas panthers helped increase fitness by reducing the presence of deleterious alleles and increasing the population size (Hedrick, 1995). While introducing other individuals can help in the short term, it’s best if the technique has positive long-term effects, and does not introduce new deleterious alleles.
You don’t want to set a ticking timebomb.
A group of researchers set out to characterize the impact of the 1995 management decision 27 years later (Ochoa et al., 2022). They confirmed that the panthers in regions that were allowed to interbreed with pumas from other regions had higher rates of heterozygosity (Figure 3). This means that their standing genetic variation is higher, and that the Florida panthers have gotten past those initial inbreeding health issues. However, the hybrid individuals are now heterozygous for health issues that were present in the Texan population.
While not a perfect scenario, additional time for the Florida panther has been bought, trading immediate health concerns for potential future ones. However, if the population can maintain a large effective population size, then there might not be strong effects. Although, if the population does decline once more, there might be an unfortunate conclusion to this story.
References
Hedrick, PW (1995) Gene Flow and Genetic Restoration: The Florida Panther as a Case Study. Conservation Biology. 9(5): 996–1007.
Hendricks S, Anderson EC, Antao T, Bernatchez L, Forester BR, Garner B, Hand BK, Hohenlohe PA, Kardos M, Koop B, Sethuraman A, Waples RS, & Luikart G (2018). Recent advances in conservation and population genomics data analysis. Evolutionary Applications. 11(8): 1197–1211.
Ochoa A, Onorato, DP, Roelke-Parker ME, Culver M, & Fitak RR (2022). Give and take: Effects of genetic admixture on mutation load in endangered Florida panthers. Journal of Heredity. 113(5): 491–499.